Введение
Напочвенные беспозвоночные играют важную роль в обеспечении устойчивости экосистем, отражают большое количество экологических факторов и выступают в качестве чувствительных биоиндикаторов природных условий и антропогенного воздействия [2]. Тем не менее, количественные сведения о напочвенных беспозвоночных урбанизированных территорий остаются фрагментарными, а закономерности их дифференциации остаются неизученными [4, 7].
Ярус животного населения может пониматься в узком смысле (s.s.) — адаптационном — и в широком смысле (s.l.) — биоценотическом [8]. В отечественной литературе напочвенная мезофауна или мезогерпетобий рассматривается в двух аспектах: как беспозвоночные определенной жизненной формы, адаптированные к обитанию на поверхности почвы, и как все беспозвоночные, фактически обнаруживаемые на поверхности почвы. В зоологических работах представлен как первый [5, 6], так и второй подход [4, 7].
Целью данной работы был анализ населения напочвенных беспозвоночных на урбанизированной территории.
Материал и методы
Материалом послужили сборы 2003—2011 гг. при помощи ловушек Барбера с использованием 4%-го формалина в качестве фиксатора и выборкой один раз в две недели.
В качестве пробных площадей исследованы относительно крупные участки, где ожидается существование комплексов беспозвоночных и где технически возможен их учет принятым методом. В зависимости от растительности и использования они объединены в несколько типов. 1) лесные биотопы — местообитания с высокой сомкнутостью древостоя и преобладанием стихийного формирования растительности; 2) дворы — биотопы, сочетающие фрагменты древесной и травянистой растительности в окружении застройки; 3) сады (садово-огородные, дачные, приусадебные, учебно-опытные участки); 4) луга — биотопы с преобладанием многолетних мезофильных и гигрофильных трав. Всего было обработано 34 выборки. В качестве факторов организации населения жужелиц исследовались сомкнутость древостоя (0 — до 10%, 1 — 10–30%, 2 — 30–70%, 3 — 70–100%); геотопология (плакор, склон и дно овражно-балочных систем); сельскохозяйственная обработка почвы; наличие застройки и искусственных покрытий на пробной площади. Характеристики пробных площадей даны в таблице 1.
При анализе герпетобионтного населения в широком (биоценотическом) смысле учитывалось обилие дождевых червей (Lumbricidae), мокриц (Isopoda, Oniscidea), пауков (Aranei), сенокосцев (Opiliones), многоножек на уровне отряда, тараканов (Blattoptera), уховерток (Dermaptera), прямокрылых (Orthoptera), клопов (Heteroptera), цикад (Auchenorrhyncha), жуков на уровне семейства. В узком (адаптационном) смысле в состав герпетобия включались мокрицы, пауки, сенокосцы, тараканы, уховертки, клопы, жуки жужелицы (Carabidae), стафилиниды (Staphylinidae), чернотелки (Tenebrionidae), мертвоеды (Silphidae), пилюльщики (Byrrhidae). Комплекс герпетобионтов описывался по уловистости (число особей на 100 ловушко-суток).
Для анализа использована среднегодовая уловистость таксонов за период с начала мая по конец октября. Для классификации комплексов напочвенной мезофауны был применен агломеративный кластерный анализ. В качестве метрики использован коэффициент Жаккара с учетом обилия. Качество формальных и априорных классификаций оценивалось по проценту ошибочной дискриминации, значению лямбда Уилкса и многомерного F-критерия в процедуре дискриминантного анализа.
Таблица 1 — Характеристика пробных площадей
|
Название биотопа |
Тип |
Сомкнутость древостоя |
Застройка |
Геотопология |
|
|
Д1 |
двор больницы № 4 |
двор |
3 |
да |
плакор |
|
Д2 |
ОРТПЦ |
двор |
2 |
да |
плакор |
|
С2 |
ЭБЦУ07 |
двор |
2 |
да |
плакор |
|
Д3.1 |
двор ЭБЦУ 06 |
двор |
2 |
да |
плакор |
|
Д3 |
дворЭБЦУ03 |
двор |
2 |
да |
плакор |
|
Д4 |
ДворЭБЦУ04 |
двор |
2 |
да |
плакор |
|
С1.2 |
с/х ЭБЦУ06 |
двор |
2 |
да |
плакор |
|
С1 |
с\х ЭБЦУ03 |
двор |
2 |
да |
плакор |
|
С1.1 |
С/х ЭБЦУ04 |
двор |
2 |
да |
плакор |
|
С4 |
Подгорная07 |
сад |
2 |
нет |
склон |
|
С5 |
Подгорная06 |
сад |
2 |
нет |
склон |
|
Л4(м) |
Жировский 03 |
лес |
3 |
нет |
дно |
|
Л7(м) |
Парк КиО |
лес |
3 |
нет |
склон |
|
Л10(м) |
Березуй03 |
лес |
2 |
нет |
дно |
|
Л11(м) |
Березуй04 |
лес |
3 |
нет |
склон |
|
Л14(м) |
Комс. роща |
лес |
3 |
нет |
склон |
|
О(г)4 |
газон |
луг |
1 |
нет |
плакор |
|
О3 |
Гр.шоссе_склон |
луг |
0 |
нет |
плакор |
|
О2 |
Гр.шоссе_низина |
луг |
1 |
нет |
плакор |
|
О1 |
Дубрава луг 2 |
луг |
1 |
нет |
плакор |
|
О9 |
Дубрава луг 1 |
луг |
1 |
нет |
плакор |
|
О2.1 |
Гр. шоссе04 |
луг |
1 |
нет |
плакор |
|
Л1 |
шир. лес |
лес |
3 |
нет |
склон |
|
С6 |
Ольговский |
сад |
2 |
нет |
плакор |
|
С7 |
Ветеран |
сад |
2 |
нет |
плакор |
|
С8 |
Северный |
сад |
2 |
нет |
плакор |
|
С9 |
Подгорная09 |
сад |
2 |
нет |
склон |
|
Л15(м) |
Жировский овраг 11 |
лес |
2 |
нет |
дно |
|
Д5(м) |
КГУ древ.сад |
двор |
3 |
да |
плакор |
|
Д(г)6 |
КГУ трав.двор |
двор |
1 |
да |
плакор |
|
С3 |
ЭБЦУ2011 |
двор |
2 |
да |
плакор |
|
О5 |
База_глина |
луг |
0 |
нет |
плакор |
|
О6 |
База_пашня |
луг |
0 |
нет |
плакор |
|
Л16(бр) |
База_березняк |
лес |
2 |
нет |
плакор |
Результаты и обсуждение
Состав населения напочвенных беспозвоночных
Основу населения напочвенных беспозвоночных во всех биотопах Калуги составляют жуки жужелицы (Coleoptera, Carabidae), стафилиниды (Staphylinidae) и пауки (Aranei); обилие прочих таксонов существенно варьирует в зависимости от типа биотопа (табл. 2).
Таблица 2 — Характеристика сообществ напочвенных беспозвоночных в соответствии с типом биотопа
| Таксон |
двор (N=12) |
сад (N=6) |
лес (N=8) |
луг (N=7) |
|||||
|
M |
SE |
M |
SE |
M |
SE |
M |
SE |
p |
|
|
Lumbricidae |
4,99 |
0,80 |
6,09 |
1,13 |
6,14 |
0,98 |
1,92 |
1,05 |
0,0250 |
|
Aranei |
25,43 |
6,60 |
46,63 |
9,33 |
34,86 |
8,08 |
66,88 |
8,64 |
0,0056 |
|
Opiliones |
18,31 |
3,45 |
13,00 |
4,88 |
22,87 |
4,22 |
7,27 |
4,51 |
0,0878 |
|
Isopoda |
26,25 |
5,83 |
24,35 |
8,24 |
13,54 |
7,14 |
27,25 |
7,63 |
0,5033 |
|
Julida |
13,28 |
2,44 |
17,47 |
3,45 |
6,34 |
2,99 |
1,51 |
3,20 |
0,0064 |
|
Polydesmida |
4,54 |
1,45 |
4,41 |
2,05 |
2,76 |
1,77 |
0,74 |
1,90 |
0,4153 |
|
Lithobiomorpha |
5,83 |
1,31 |
5,19 |
1,86 |
2,63 |
1,61 |
2,43 |
1,72 |
0,3008 |
|
Geophilomorpha |
0,22 |
0,20 |
0,20 |
0,28 |
0,33 |
0,24 |
1,01 |
0,26 |
0,0970 |
|
Larva insectae |
18,43 |
2,34 |
22,18 |
3,31 |
17,57 |
2,86 |
11,20 |
3,06 |
0,1180 |
|
Dermaptera |
9,80 |
2,64 |
4,58 |
3,73 |
5,05 |
3,23 |
2,04 |
3,46 |
0,3252 |
|
Orthoptera |
0,40 |
0,93 |
0,88 |
1,31 |
0,14 |
1,14 |
9,85 |
1,22 |
0,0000 |
|
Auchenorrhyncha |
6,40 |
1,80 |
4,94 |
2,54 |
3,35 |
2,20 |
17,82 |
2,35 |
0,0005 |
|
Heteroptera |
2,87 |
0,68 |
1,38 |
0,97 |
2,40 |
0,84 |
4,26 |
0,90 |
0,1926 |
|
Coleoptera, в т. ч. |
126,99 |
16,04 |
142,99 |
22,69 |
159,83 |
19,65 |
164,08 |
21,01 |
0,4601 |
|
Carabidae |
50,73 |
6,02 |
72,14 |
8,52 |
45,92 |
7,38 |
37,27 |
7,89 |
0,0372 |
|
Staphylinidae |
32,88 |
5,40 |
29,01 |
7,64 |
44,66 |
6,62 |
50,67 |
7,07 |
0,1158 |
|
Silphidae |
2,52 |
1,82 |
2,08 |
2,57 |
7,04 |
2,23 |
2,30 |
2,38 |
0,3550 |
|
Elateridae |
1,41 |
0,44 |
2,67 |
0,62 |
0,92 |
0,53 |
5,42 |
0,57 |
0,0000 |
|
Byrrhidae |
0,45 |
0,25 |
0,22 |
0,35 |
0,19 |
0,30 |
1,76 |
0,33 |
0,0049 |
|
Nitidulidae |
1,96 |
0,28 |
0,62 |
0,40 |
0,60 |
0,34 |
0,63 |
0,37 |
0,0069 |
|
Catopidae |
21,82 |
6,19 |
11,77 |
8,76 |
30,13 |
7,58 |
2,02 |
8,11 |
0,0843 |
|
Curculionidae |
10,15 |
8,05 |
4,91 |
11,39 |
26,89 |
9,86 |
43,26 |
10,54 |
0,0536 |
|
Chrysomelidae |
2,24 |
1,19 |
15,94 |
1,68 |
1,46 |
1,45 |
6,28 |
1,55 |
0,0000 |
|
Tenebrionidae |
— |
— |
— |
— |
0,02 |
4,03 |
10,24 |
4,31 |
0,2408 |
Примечания:
M — среднее арифметическое значение уловистости (экз./100 ловушко-суток),
SE — стандартное отклонение уловистости,
p — уровень значимости межгрупповых различий по F-критерию (значимые p выделены полужирным шрифтом)
Классификация городских биотопов по населению беспозвоночных
Герпетобий в широком смысле. В целом по населению напочвенных беспозвоночных можно противопоставить биотопы с преобладанием древесной растительности и луговые биотопы (рис. 1, 2). Луговые биотопы отличаются высоким обилием пауков, цикад, клопов, низким обилием диплопод, костянок, катопид. Среди биотопов лесного типа два оказались ближе к лугам: влажный участок днища оврага (Л15) и березняк на пустыре в окружении луговой растительности (Л16). Садовые участки отличаются от других биотопов с древесной растительностью повышенной уловистостью пауков, жужелиц, щелкунов, листоедов. Дворы совершенно не дискриминируются от лесных биотопов (рис.).
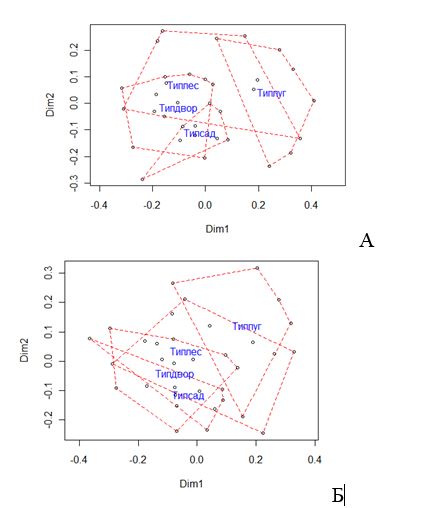
Рисунок 1 — Ординация комплексов напочвенных беспозвоночных в пространстве двух осей многомерного неметрического шкалирования. Пунктиром показаны границы типов биотопов. А — герпетобий в широком смысле, Б — герпетобий в узком смысле.
В свою очередь, луга по населению беспозвоночных неоднородны и могут быть разделены на два кластера (рис. 2, 3). «Сухо-луговой» кластер включает газон вдоль автомагистрали Грабцевское шоссе и три смежных биотопа на зарастающем пустыре в районе «База», в т. ч. молодой березняк. Эти биотопы отличаются высоким обилием геофилов, пилюльщиков, долгоносиков. «Влажно-луговой» кластер (участки разнотравно-злаковой растительности, а также участок дна оврага) отличается от предыдущего высокой уловистостью стафилинид, мертвоедов и мокриц, меньшей уловистостью жужелиц и долгоносиков.
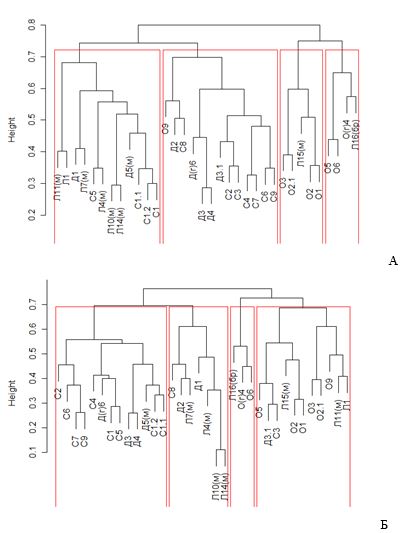
Рисунок 2 — Результаты кластерного анализа населения напочвенных беспозвоночных Калуги (индекс Жаккара с учетом обилия, метод максимальной связи). А — герпетобий в широком смысле, Б — герпетобий в узком смысле. Обозначения пробных площадей см. в табл. 1.
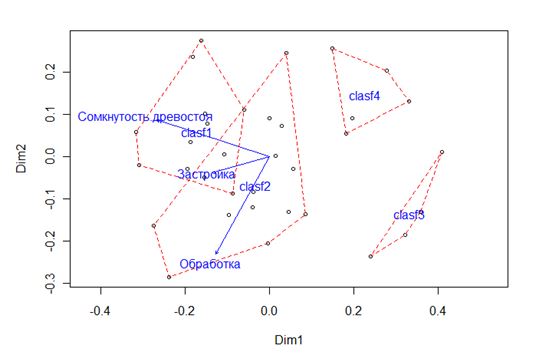
Рисунок 3 — Ординация кластеров сообществ напочвенных беспозвоночных в пространстве двух осей многомерного неметрического шкалирования.
Пунктиром показаны границы кластеров формальной классификации (clasf1-4). Показатели корреляции с осями: сомкнутость.древостоя (r2=0,588, p=0,001), застройка (r2=0,148, p=0,081), обработка (r2=0,116, p=0,159).
По сравнению с жуками-жужелицами, изученными нами в тех же биотопах [1], население напочвенных биотопов более изменчиво в биотопах с древесной растительностью. Кластер луговых биотопов для напочвенных беспозвоночных более компактен, чем для жужелиц. Промежуточное положение садов между лесными и луговыми биотопами по напочвенным беспозвоночным не столь отчетливо, как по жужелицам.
Герпетобий в узком смысле. При исключении из анализа фитофильных, подстилочных и геобионтных таксонов была получена плохо интерпретируемая классификация, дающая к тому же ошибочное распознавание отдельных образцов при дискриминантном анализе (рис. 1, 2). По результатам дискриминантного анализа данная классификация значима только для пауков. Значимо отделен от прочих типов сообществ 4-й кластер (примерно соответствует «сухо-луговому» типу предыдущей классификации) и 3-й кластер (влажно-луговой с добавлением лесов). 1-й кластер, включающий преимущественно древесные насаждения, выделяется по низкому обилию мокриц и пауков, однако очень слабо дифференцирован от второго.
С формальных позиций классификация, основанная на населении герпетобия в биоценотическом смысле, гораздо качественнее, чем классификация, основанная на населении герпетобия в узком смысле. При этом существенных различий между классификациями с учетом уловистости и классификациями по долям нет (табл. 3).
Таблица 3 — Характеристики классификаций герпетобия по результатам дискриминантного анализа
|
Классификация |
Число классов |
Лямбда Уилкса |
F |
p |
Число случаев ошибочной дискриминации |
|
Формальные классификации: |
|||||
|
по уловистости герпетобия s.l. |
4 |
0,00001 |
10,689 |
< 0,0001 |
0 |
|
по % герпетобия s.l. |
4 |
0,00005 |
9,3409 |
< 0,0001 |
0 |
|
по уловистости герпетобия s.s. |
4 |
0,01836 |
3,4715 |
<0,0001 |
1 |
|
по % герпетобия s.s. |
4 |
0,01385 |
3,9392 |
<0,0001 |
3 |
|
Априорные классификации: |
|||||
|
по типу биотопа |
4 |
0,00022 |
4,1121 |
0,0006 |
1 |
|
по сомкнутости древостоя |
4 |
0,00009 |
6,9327 |
< 0,0001 |
0 |
Факторы дифференциации населения напочвенных беспозвоночных
Сомкнутость древостоя можно считать ведущим фактором дифференциации населения беспозвоночных (рис. 3). Она определяет обилие 12 таксонов мезогерпетобия (табл. 3.10). Обилие прямокрылых, цикад, клопов снижается при возрастании сомкнутости древостоя. Пауки и жуки щелкуны избегают биотопов с преобладанием древесной растительности. Листоеды значимо избегают только биотопы с наиболее плотным древостоем (3). Геофилы и пилюльщики предпочитали биотопы без деревьев. Кивсяки предпочитают биотопы с преобладанием древесной растительности (2 и 3). Сенокосцы и жуки катопиды тяготеют к биотопам наиболее высокой степени сомкнутости.
Данные материалы подтверждают доказанную в экспериментах ведущую роль растительности в дифференциации мезофауны [3]. Конкретные причины предпочтения древесной и травянистой растительности для разных таксонов неодинаковы, в частности, доказана роль подстилки для распределения беспозвоночных [9, 10].
В то же время не всегда по сомкнутости древостоя можно предсказать население напочвенных беспозвоночных, что подтверждает мнение некоторых исследователей, работавших на малонарушенных территориях [5]. Так, травянистый двор КГУ обладает таким же животным населением, как и окружающие его деревянистые биотопы, а березняк в районе «База» мало отличается от окружающих лугов. В то же время соседние биотопы могут контрастировать по животному населению (напр., газон и луговая растительность на Грабцевском шоссе). Вероятно, перечисленные случаи связаны не столько с прямым влиянием окружающих биотопов, сколько с феноменом «экологической памяти» герпетобия [6].
Обработка почвы значимо повышает уловистость кивсяков, жужелиц, листоедов, снижает уловистость долгоносиков.
Значимость наличия искусственных покрытий удалось показать лишь для одного таксона — стафилинид. Эти жуки избегают стаций с искусственными покрытиями. В биотопах с высоким процентом запечатанности низка уловистость жесткокрылых в целом.
Тип местоположения (геотопология) участка значим только для пауков, которые предпочитают дно овражно-балочной сети склонам и плакорным участкам, и долгоносиков, избегающих дна балок.
Сообщества в биотопах, окруженных многоэтажной застройкой, не отличаются значимо от сообществ в биотопах, не находящихся в прямом контакте с застройкой, ни по одному таксону.
Заключение
Таким образом, население напочвенных беспозвоночных, рассматриваемое на уровне таксонов высокого ранга, достаточно хорошо дифференцируется на комплексы, интерпретируемые по признакам биотопа.
Дифференциация комплексов напочвенной мезофауны на урбанизированной территории осуществляется по градиентам развития древесной и травянистой растительности. На обилие отдельных таксонов влияет обработка почвы, геотопология и искусственные покрытия.
Исключение из анализа таксонов, не входящих в герпетобий в узком смысле, существенно ухудшает качество и интерпретируемость классификации.
Литература
- Алексанов В. В., Алексеев С. К., Сионова М. Н. Типология комплексов жужелиц урбанизированного ландшафта (на примере города Калуги) // Проблемы региональной экологии. 2012. № 6. С. 99–109.
- Богач Я., Ружечка В. Анализ экологических групп видов сообществ напочвенных беспозвоночных как показатель качества окружающей среды // Экология. 1988. № 6. — С. 59 — 61.
- Бызова Ю. Б. О скорости образования и дифференцировки комплексов беспозвоночных в искусственных ценозах // Проблемы почвенной зоологии. — Тбилиси: Мецниерба, 1987. — С. 51–53
- Еремеева Н. И. Структура и экологические механизмы формирования мезофауны членистоногих урбанизированных территорий (на примере г. Кемерово): Автореф. дис. … д-ра биол. наук. — Новосибирск, 2006. — 42 с.
- Майманакова И. Л. Роль лесных и степных экосистем Минусинской котловины в формировании комплексов жесткокрылых-герпетобионтов: Автореф. дис. … канд. биол. наук. Красноярск, 2009. 22 с.
- Мордкович В. Г. Зоологическая характеристика примитивных почв в осушных зонах соленых озер юга Сибири // Зоологический журнал. 1973. Т. LII. Вып. 9. С. 1321–1329.
- Савосин Н. А. Эколого-фаунистическая характеристика герпетобионтного населения членистоногих крупного промышленного центра: Автореф. дис. … канд. биол. наук. — Барнаул, 2010. — 23 с.
- Чернов Ю. И. Понятие «животное население» и принципы геозоологических исследований // Журнал общей биологии. — 1971. — Т. 32. — Вып. 4. — С. 425–438.
- Kazemi F., Beecham S., Gibbs J., Clay R. Factors affecting terrestrial invertebrate diversity in bioretention basins in an Australian urban environment // Landscape and Urban Planning. 2009. vol. 92. P. 304–313
- McIntyre N. E. The ecology of urban arthropods: a review and a call to action // Annals of the Entomological Society of America. 2000. 93. P. 825–835.