Введение
Использование
клеточных культур растений в качестве источника ценных метаболитов может стать
реальной альтернативой применения интактным растениям, что обусловлено, в
первую очередь, экологичностью такого производства, независимостью культивирования
от климатических условий и вредителей, автоматизацией производственного
процесса [5]. Пажитник греческий (Trigonella
В этой связи целью представленной работы явилось исследование динамики роста и накопления фенольных соединений в ходе ростового цикла суспензионной культуры пажитника греческого.
Материал и методы исследования
Объектом
изучения служила суспензионная культура пажитника греческого, инициированная из
гетеротрофного каллуса листового происхождения пажитника ярового сорта Ovari 4 [8].
Для инициации суспензионной культуры
помещали 6–7 г свежей
рыхлой массы каллусных клеток в колбу на 500 мл с 200 мл стерильной
жидкой питательной среды
[3], соответствующей по составу среде, используемой для выращивания данной
каллусной ткани. Среду, минеральная основа которой соответствовала среде
Мурасиге и Скуга, дополняли регуляторами роста: 1,0 мг/л 2,
Определение
ростовых и биосинтетических показателей суспензионной культуры пажитника
греческого проводили на
Измерения проводили на протяжении трех пассажей. Все результаты обработаны статистически. Данные на графиках представлены в виде средних значений ± стандартная ошибка средней.
Результаты исследования и их обсуждение
Одной из важнейших характеристик роста клеточных
культур, позволяющих оценить активность ростовых процессов на разных этапах
выращивания и определить ее оптимальную продолжительность, является кривая
ростового цикла. Из графика,
представленного на рис. 1, видно, что латентная фаза роста суспензионной
культуры пажитника греческого длилась до
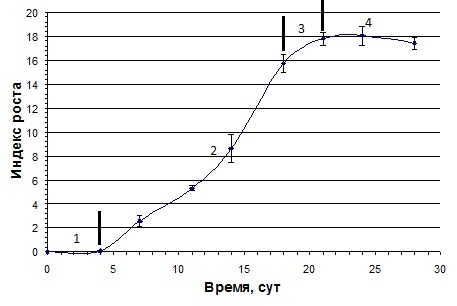
Рисунок 1 — Кривая роста суспензионной культуры пажитника греческого. Фазы роста: 1 — латентная, 2 — логарифмическая, 3 — замедленная, 4 — стационарная
Кроме изучения динамики накопления биомассы, важным этапом исследования суспензионных культур является описание их морфофизиологических особенностей, в частности изменение степени их агрегированности, жизнеспособности и морфологии в ходе ростового цикла.
Из
диаграмм, представленных на рис. 2, видно, что на протяжении роста
суспензионной культуры преобладали одиночные клетки и мелкие агрегаты.
Процентное содержание первых от общего числа фракций составляло 56%
в начале ростового цикла (
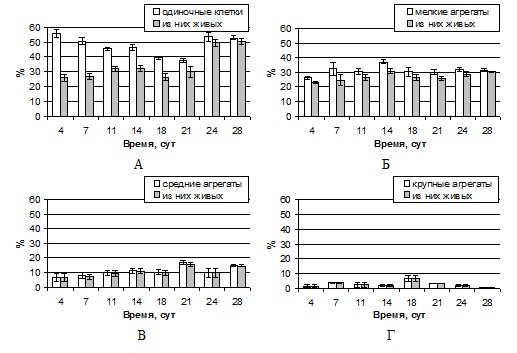
Рисунок 2 — Процентное содержание различных фракций клеток в ходе ростового цикла суспензионной культуры пажитника греческого: А — одиночные клетки, Б — мелкие агрегаты, В — средние агрегаты, Г — крупные агрегаты
Анализ
жизнеспособности культуры показал, что количество живых одиночных клеток в ходе
латентной фазы (
Изучение морфологии суспендированных клеток пажитника греческого показало, что в культуре преобладали клетки округлой и неправильной формы, часто находящиеся в скоплениях по 6—8 клеток (рис. 3). Также встречались одиночные крупные клетки червеобразной, круглой, спиралевидной, овальной и бобовидной формы. Необходимо отметить, что перечисленные выше типы клеток присутствовали в суспензии клеток на всех этапах ростового цикла.
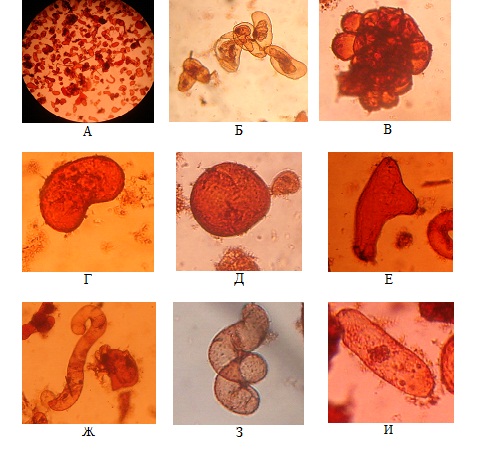
Рисунок 3 — Морфология суспендированных клеток пажитника греческого: А — общий вид окрашенных клеток; Б — скопление клеток овальной и неправильной формы; В — средний агрегат, состоящий из клеток округлой формы; Г — бобовидная клетка; Д — гигантская круглая клетка; Е — клетка неправильной формы; Ж — червеобразная клетка; З — спиралевидная клетка; И — клетка овальной формы
Представленные
результаты свидетельствуют о том, что ростовая кривая суспензионной культуры
пажитника греческого имеет стандартную
Ранее нами было показано, что основу полифенольного комплекса каллусных культур пажитника греческого составляют фенолкарбоновые кислоты [9]. Мы предположили, что суспензионная культура, инициированная из каллусной ткани пажитника греческого, может сохранить способность к преимущественному накоплению фенолокислот. В этой связи следующим этапом нашего исследования было определение динамики изменения общего содержания соединений фенольной природы и в частности фенолкарбоновых кислот в суспензии клеток пажитника греческого, и выявление связи между указанными биохимическими характеристиками.
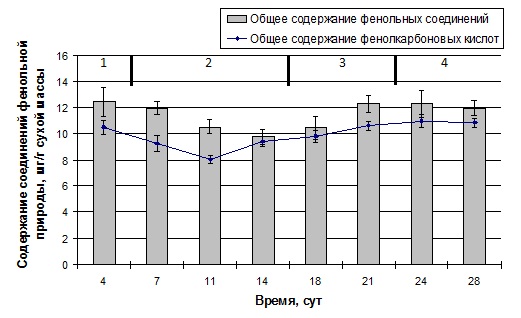
Рисунок 4 — Динамика накопления фенольных соединений и фенолкарбоновых кислот в ходе ростового цикла суспензионной культурой пажитника греческого. Фазы роста: 1 — латентная, 2 — логарифмическая, 3 — замедления, 4 — стационарная
Показано,
что общее содержание фенольных соединений в ходе латентной и начальной логарифмической фазах роста суспензионной
культуры изменялось незначительно (рис. 4). К
Исходя
из вышепредставленных данных, можно заключить, что в суспензии клеток пажитника
греческого фенольные соединения накапливаются на протяжении всего ростового
цикла, причем нет достоверных различий между содержанием данных метаболитов на
начальных этапах ростового цикла (
Анализ изменения содержания фенольных соединений и фенокарбоновых кислот в суспензии клеток пажитника греческого в ходе ее ростового цикла позволяет судить о наличии положительной связи между данными биохимическими показателями. Для оценки взаимосвязи был определен коэффициент корреляции Пирсона. Установлена прямая, сильная и достоверная связь между исследуемыми параметрами: rху = 0,66, р > 95%. Так, с вероятностью безошибочного прогноза больше 95% характер изменения содержания фенолкарбоновых кислот в суспензионной культуре пажитника греческого на разных стадиях роста соответствует характеру динамики накопления фенольных соединений в целом. Таким образом, в изучаемой нами суспензии клеток на всех этапах ее ростового цикла именно фенолкарбоновые кислоты составляют основную долю полифенольного комплекса.
Заключение
Представленные результаты свидетельствуют о том, что суспензионная культура пажитника греческого характеризуется высокой жизнеспособностью и продуктивностью по биомассе, а также способностью на всём протяжении ростового цикла синтезировать соединения фенольной природы, основную долю которых составляют фенолкарбоновые кислоты. Это позволяет рассматривать ее как перспективный альтернативный источник получения физиологически активных веществ фенольной природы в промышленных масштабах.
Работа
выполнена при финансировании Белорусским республиканским фондом фундаментальных
исследований (проект № Б13
Список литературы
- Гаврилин М. В.,
Попова О. И., Губанова
Е. А. Фенольные соединения надземной части шалфея мускатного (Salvia sclarea L.), культивируемого в Ставропольском крае // Химия растительного сырья. 2010. Вып. 4. — С. 99–104. - Георгиевский
В. П., Комиссаренко П. Ф., Дмитрук
С. Е. Биологически активные вещества лекарственных растений / Новосибирск. Наука, Сиб.отд-ние . 1990. — 333 с. - Дитченко
Т. И. Культура клеток, тканей и органов растений: методические рекомендации к лабораторным занятиям / Мн. БГУ. 2007. — 22 c. - Калинин Ф. Л., Методы культуры тканей в физиологии и биохимии растений / Киев. Наук. думка. 1980. — 448 c.
- Chawla
H. S. Introduction to plant biotechnology (2nd ed.) / Enfield. Science Publishers. 2002. — 538 p. Godoy-Hernandez G.,Vazquez-Flota F. A. Growth measurements: estimation of cell division and cell expansion // Plant Cell Culture Protocols. Series: Methods in Molecular Biology. 2006.v. 318 (2). — P. 51–58.- Kaviarasan S., Naik
G. H., Gangabhagirathi R., Anuradha C.
v. , PriyadarsiniK. I. In vitro studies on antiradical and antioxidant activities of fenugreek (Trigonellafoenum-graecum ) seeds // Food Chemistry. 2007.v. 103. — P. 31–37. - Lohvina H. O.,
Makai S., Ditchenko T. I., Reshetnikov
v. N., Spiridovich E.v. , Yurinv. M. Induction of callus from leaves and stems of Trigonellafoenum-graecum varieties // Acta Agronomica Óváriensis. 2012.v. 54 (2). — P. 29–37. - Lohvina H. O.,
Yurin
v. M. Phenolics and antioxidant potential of fenugreek cell cultures // Book of Abstracts of The 17th International Pushchino School Conference of Young Scientist «Biology — The Science of The XXI Century», Pushchino, Russia. April21-26th . 2013. — P. 384–385. - Slinkard K., Singleton
v. L. Total phenol analysis: automation and comparison with manuel methods // American journal of enology and viticulture. 1977.v. 28. — P. 49–55.