Введение
Бешенство является опасным нейроинфекционным заболеванием теплокровных животных (в том числе и человека), которое встречается более чем в 150 странах мира [7, 10]. Возбудителем данного заболевания является РНК-содержащий вирус рода Lyssavirus, семейства Rhabdoviridae, порядка Mononegavirales. Заражение бешенством главным образом происходит через укус, в ходе которого инфекция передаётся со слюной. Основной мишенью вируса бешенства является центральная нервная система (ЦНС). Вирус бешенства является единственным из царства Virae, который поражает всех теплокровных животных с летальностью 100 %. От укусов животных, больных бешенством, ежегодно в мире погибает более 55 000 человек. В связи с этим согласно оценке Всемирной Организации Здравоохранения (ВОЗ) бешенство входит в пятёрку болезней, которые наносят наибольший ущерб человечеству и мировой экономике. Единственным методом борьбы с данным заболеванием являются пред- и постэкспозиционная профилактика, в рамках которой используются антирабические иммуноглобулин (АИГ) и вакцина (в дальнейшем — антирабические препараты) [1, 7, 10, 11].
Для получения антирабических препаратов и контроля их качества используются аттенуированные (или фиксированные) штаммы вируса бешенства, которые получают в ходе воздействия на полевые изоляты мутагенов или других факторов, позволяющих менять свойства вирионов. В отличие от полевых изолятов штаммы фиксированного вируса бешенства могут быть полностью или частично лишены способности к заражению целого организма животного или человека, но сохраняют возможность заражать ткани и клетки при непосредственном контакте. Наиболее распространёнными в производстве антирабических препаратов являются фиксированные штаммы вируса бешенства L. Pasteur и CVS, выделенные более 100 лет назад из полевых изолятов, ослабленные путём пассирования через мозг кроликов и адаптированные к мозгу мышей и клеточным линиям. С 1996 года ВОЗ рекомендует при производстве антирабических препаратов полностью отказаться от использования вируса бешенства, полученного пассированием через мозговую ткань животных, и использовать для накопления вирусной суспензии клеточные культуры [8, 11].
ПАО «Фармстандарт-Биолек» (Харьков, Украина) занимается разработкой антигена вируса бешенства для дальнейшего получения АИГ и антирабических вакцин для человека и животных с использованием постоянных клеточных линий и фиксированных штаммов L. Pasteur и CVS. Технологическая схема производства антирабических препаратов требует долгосрочного хранения промышленных штаммов вируса бешенства в виде суспензии. Большинство исследователей рекомендуют хранить вирусную суспензию при различных низких температурах (-20, -60, -70, -80°С) или в лиофилизированном состоянии [4, 9, 11, 12]. Но метод лиофилизации не подходит для производства, так как требует дополнительного пассирования и позволяет хранить только небольшие объёмы, которых недостаточно для производственных целей. В связи с этим в условиях производства необходимо хранить вирусную суспензию при низких температурах. Однако данные по эффективности низкотемпературного хранения аттенуированных штаммов вируса бешенства в суспензии противоречивы, что ставит под угрозу качество антирабических препаратов, полученных с использованием суспензии вируса бешенства, хранившейся при низких температурах.
Цель исследования
Цель исследования — экспериментально определить эффективный температурный режим для долгосрочного хранения суспензии вируса бешенства штаммов L. Pasteur и CVS в условиях производства.
Материалы и методы
В качестве субстрата для накопления суспензии вируса бешенства использовали перевиваемую клеточную линию ВНК-21 / C13 (ATCC CCL-10), которая широко применяется для производства культуральной вакцины против бешенства [5, 6, 8, 12]. Клеточную линию накапливали в пристеночном монослое 1 сутки в стерильных пластиковых флаконах (SPL, Германия). В ходе исследования использовали фиксированные штаммы вируса бешенства L. Pasteur (адаптированный к клеточной линии Vero, № 2061 / Vero, 15 пассаж) и CVS (challenge virus standard), рекомендованные ВОЗ для производства АИГ и антирабических вакцин для животных и человека [8, 12]. Штамм L. Pasteur был адаптирован к клеточной линии ВНК-21 / C13 [6] и предоставлен Институтом Пастера (Нови-Сад, Сербия) вместе с технологическим регламентом производства антирабического антигена.
Вирусную суспензию культивировали в течении 4-х суток с момента заражения в поддерживающей среде DMEM (Dulbecco's Modified Eagle's Medium) c 0, 2 % бычьего альбумина (PAA, Австрия) в СО2-инкубаторе (Binder, Германия) при 33ºС и 5 % СО2. Сбор вирусной суспензии проводили на 4 сутки после заражения. Полученную вирусную суспензию центрифугировали при 3000 об/мин 15 минут в рефрижераторной центрифуге (MPW, Польша) при 4ºС для удаления клеточного детрита. Супернатант собирали, добавляли к нему сахарозу с конечной концентрацией 5 %, расфасовывали в пластиковые криопробирки (PAA, Австрия) и замораживали при -20 и -80ºС в морозильных камерах (National Lab, Германия). Контроль температуры, при которой хранили криопробирки, производили с помощью спиртовых термометров [5, 6, 8, 12].
Для определения специфической инфекционной активности вирусной суспензии
использовали метод титрации вируса в культуре клеток линии
ВНК-21 / C13. Суспензию вируса бешенства размораживали через 1
неделю, 1 , 2, 3, 6 и 12 месяцев хрнения при низких температурах (срок
наблюдения) и титровали с коэффициентом разведения 5 на культуре клеток
ВНК-21 / C13 в 96-луночных планшетах (SPL, Германия) в 5-ти повторах
(n = 5). Культивировали клетки 48 часов в СО2-инкубаторе
при 37ºС и 5 % СО2, затем
фиксировали монослой клеточной лини охлаждённым при -20ºС ацетоном. Так как промышленные
штаммы вируса бешенства не оказывают на клеточные линии цитопатогенного
действия, то для определения инфекционной активности вирусной суспензии
использовали метод непрямой флуоресценции. Для этого применяли специфические
моноклональные антитела к вирусу бешенства, меченные изотиоцинатом флуоресцина
(FITC) (Fujirebio, U.S.A.), которыми производили окрашивание
клеточного монослоя. Учёт результатов производили с помощью медицинского микроскопа
(МИКМЕД-6, ЛОМО, Россия) (увеличение 100×) с люминисцентной насадкой. При
микроскопировании учитывали яркое специфическое свечение зелёного цвета,
которое свидетельствует о наличии вируса бешенства в клетках, и лунки с таким
свечением отмечали как положительные (рисунок 1) [4, 5, 9, 12].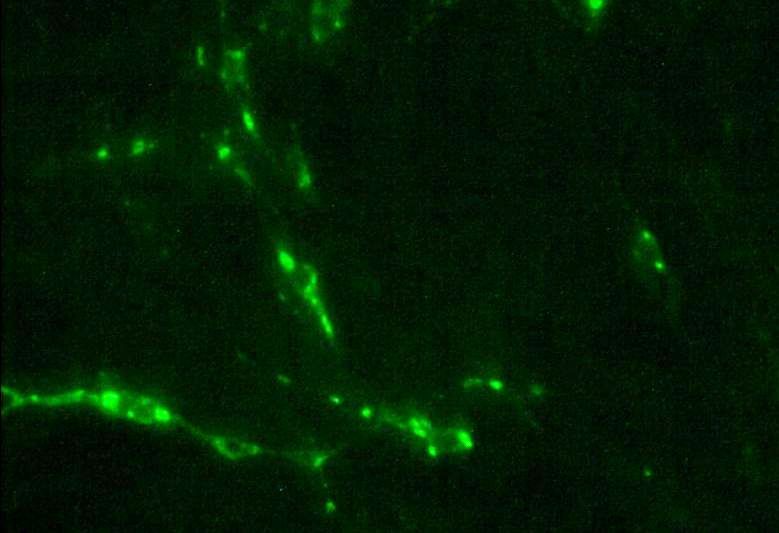
Рисунок 1 — Клетки лини ВНК-21 / С13, инфицированные вирусом бешенства штамма L. Pasteur. Окраска FITC-меченными моноклональными антителами, увеличение 100×
Расчёт титра вируса проводили с помощью формулы Спирмена-Карбера и выражали в десятичном логарифме 50 %-ой фокусформирующей инфицирующей дозы (lg FFD50):
![]() , где
, где
x0 — lg наибольшего разведения, в котором во всех лунках отмечается положительное свечение;
d — lg фактора разведения;
ni — общее количество лунок, приходящихся на каждое разведение титрации вирусной суспензии;
ri — количество положительных лунок в каждом разведении титрации вирусной суспензии [3, 9, 12].
Статистический анализ данных осуществляли с помощью стандартных пакетов компьютерных программ Microsoft Excel-2010 и Past. Вид распределения определяли с помощью W-критерия Шапиро-Уилка, достоверность различий между группами данных рассчитывали с использованием параметрического t-критерия Стьюдента (для групп с нормальным распределением данных). Расхождение считали статистически значимым при p ≤ 0, 05.
Результаты и обсуждение
В ходе проведенных исследований была проанализирована сохранность специфической инфекционной активности суспензии вируса бешенства штаммов L. Pasteur и CVS после хранения при температурах -20 и -80°С через 1 неделю, 1, 2, 3, 6 и 12 месяцев (срок наблюдения). В качестве контроля использовали данные по хранению вируса на протяжении 1-ой недели.
Штамм L. Pasteur (-20°С)
При анализе данных по хранению вирусной суспензии
промышленного штамма L. Pasteur при -20ºС были получены
следующие результаты. Активность вирусной суспензии через 1 неделю после
замораживания при температуре -20°С составила 8,11±0,29 lg FFD50 (n = 5) (рисунок 2). При сравнении данных по специфической
активности вируса после 1-ого месяца хранения различия не были статистически
значимыми (р ≥ 0, 05). После 2-х месяцев хранения при этой
температуре активность вируса данного штамма начала снижаться (р ≤ 0,
05 ). Аналогичные данные были получены при сравнении активности вирусной
суспензии после хранения на протяжении 3-х, 6-ти и 12-ти месяцев при
температуре -20°С. Различия между данными по специфической активности после
указанных сроков хранения были статистически значимы (р ≤ 0, 05).
Через 12 месяцев хранения при температуре -20°С активность вируса штамма
L. Pasteur составила 4, 75 ± 0, 19 lg FFD50, что на 3, 36 lg FFD50 меньше, чем данные контроля. 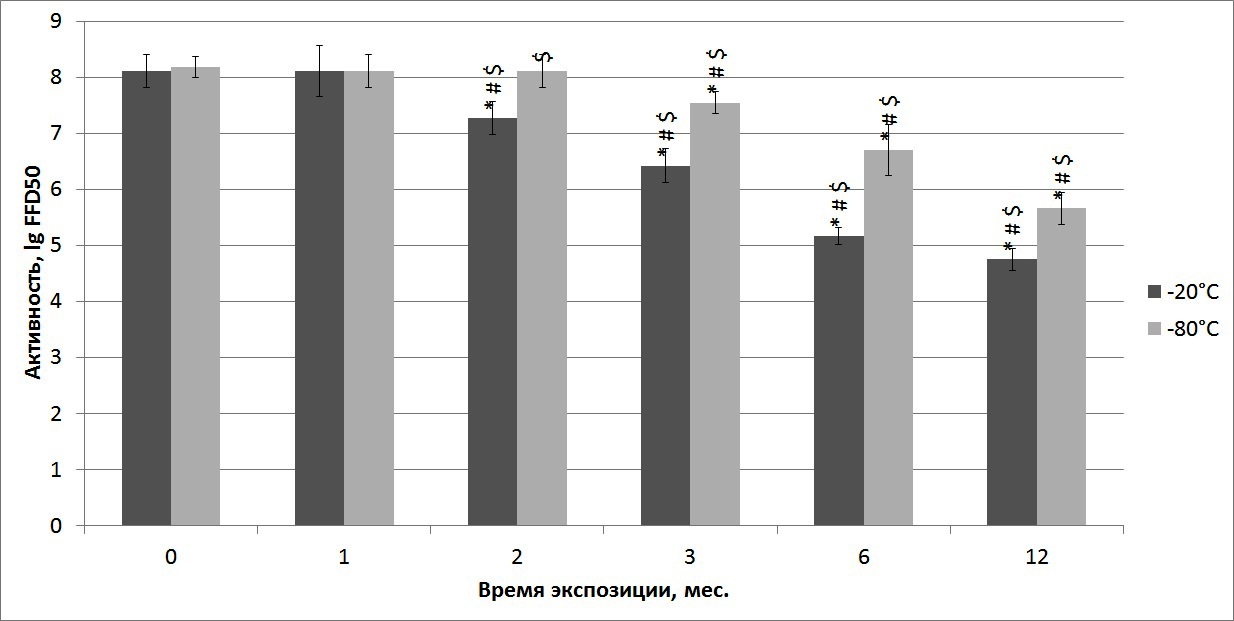
Рисунок 2 — Динамика изменения инфекционной активности вируса бешенства штамма L. Pasteur в ходе хранения при температурах -20°С и -80°С, * — статистически значимые различия между контролем и последующими сроками хранения; # — статистически значимые различия между каждым из сроков; $ — статистически значимые различия между показателями активности одного срока хранения, но при разных температурах
Штамм L. Pasteur (-80°С)
Специфическая инфекционная активность вирусной суспензии штамма L. Pasteur через 1 неделю после замораживания при температуре -80°С составила 8, 18 ± 0, 19 lg FFD50 (n = 5) и не изменялась в течение 2-х месяцев (р ≥ 0, 05). Между 2-м и 3-м месяцами хранения при температуре -80°С инфекционная активность вируса начала достоверно снижаться (р ≤ 0, 05 ), как и на протяжении последующего хранения. Через 12 месяцев хранения при температуре -80°С активность вируса штамма L. Pasteur составила 5, 66 ± 0, 29 lg FFD50, что на 2, 52 lg FFD50 меньше, чем данные контроля. Со 2-ого по 12-й месяцы хранения при -80°С показатели инфекционной активности были достоверно выше (р ≤ 0, 05 ), по сравнению с образцами, хранившимися при -20°С.
Штамм CVS (-20°С)
Активность вирусной суспензии промышленного штамма CVS через 1 неделю после хранения при температуре -20°С составила 6, 78 ± 0, 19 lg FFD50 (n = 5) (рисунок 3) и не изменялась до 2-х месяцев хранения (р ≥ 0, 05). Между 2-м и 3-м месяцами хранения при данной температуре произошло достоверное снижение (р ≤ 0, 05 ), активности, как и на протяжении последующего хранения. Через 12 месяцев хранения при температуре -20°С активность штамма CVS составила 4, 96 ± 0, 29 lg FFD50, что на 1, 82 lg FFD50 меньше, чем специфическая активность вируса при хранении в данном температурном режиме в течение недели.
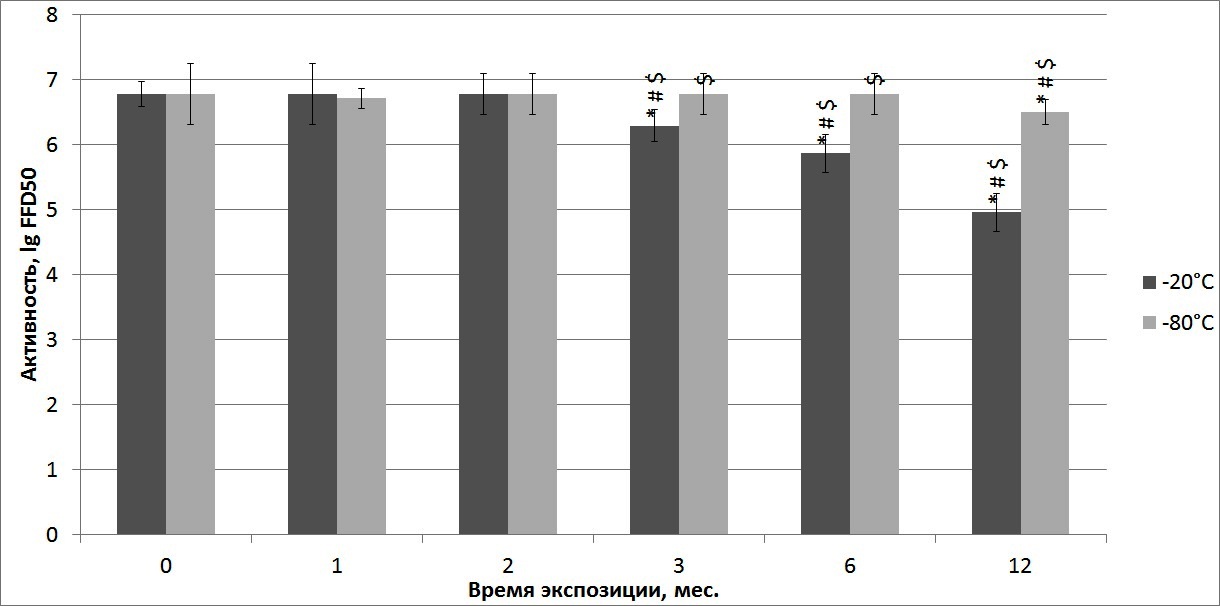
Рисунок 3 — Динамика изменения инфекционной активности вируса бешенства штамма CVS в ходе хранения при температурах -20°С и -80°С, * — статистически значимые различия между контролем и последующими сроками хранения; # — статистически значимые различия между каждым из сроков; $ — статистически значимые различия между показателями активности одного срока хранения, но при разных температурах
Штамм CVS (-80°С)
Активность вирусной суспензии штамма CVS через 1 неделю после замораживания при температуре -80°С составила 6, 78 ± 0, 47 lg FFD50 (n = 5) и не изменялась в течение 6-ти месяцев (р ≥ 0, 05). Достоверное снижение (р ≤ 0, 05) показателя активности вируса произошло между 6-м и 12-м месяцами хранения при данной температуре. После 12 месяцев хранения при температуре -80°С активность вирусной суспензии снизилась на 0,28 lg FFD50 и составила 6, 5 ± 0, 19 lg FFD50 по сравнению с данными контроля. С 3-ого по 12-й месяцы хранения при -80°С показатели инфекционной активности данного штамма были достоверно выше (р ≤ 0, 05 ), по сравнению с образцами, хранившимися при -20°С.
В результате проведенных исследований более низкие результаты сохранности обоих штаммов вируса были получены при хранении вирусной суспензии при температуре -20°С. Это может быть связано с тем, что эвтектическая температура солевых растворов, входящих в состав среды консервирования и ультраструктурных компонентов вирионов находится ниже -20°С, и вирионы подвергаются действию повреждающих факторов, связанных с кристаллизацией жидкой фазы, изменением рН и гиперконцентрацией электролитов [2].
Результаты исследования показали, что фиксированный штамм CVS более устойчив к хранению при низких температурах, чем штамм L. Pasteur. Предположительно, различия криоустойчивости изучавшихся промышленных штаммов вируса бешенства обусловлены особенностями строения их капсидов. Этот вопрос требует отдельного изучения.
Полученные результаты по сохранности вируса бешенства промышленных штаммов в суспензии не соответствуют требованиям производства, так как необходимо сохранение спицефической ативности вируса на протяжении более длительных сроков. Поэтому следует провести исследования по применению различных режимов замораживания вирусной суспензии и использованию различных криозащитных сред. Полученные данные, вероятно, смогут позволить хранить промышленные штаммы вируса бешенства при низких температурах более длительное время без потери их специфической активности.
Выводы
- В ходе проведенных исследований установили различия в криорезистентности промышленных штаммов вируса бешенства L. Pasteur и CVS. Фиксированный штамм вируса бешенства CVS дольше сохраняет свою специфическую инфекционную активность после хранения при режимах -20 и -80°С.
- Экспериментально определили, что хранение вирусной суспензии при температуре -80°С по сравнению с температурой -20°С обеспечивает более высокую сохранность специфической инфекционной активности обоих промышленных штаммов вируса бешенства.
Литература
- Львов Д. К., Алимбарова Л. М., Альховский С. В. и др. Медицинская Вирусология / Под ред. Львова Д. К. — М.: МИА, 2008. — 656 c.
- Цуцаева А. А., Микулинский Ю. Е., Высеканцев И. П. Холодовой стресс и биологические системы / Под ред. Цуцаевой А. А. — К.: Наук. думка, 1991. — 176 с.
- Aubert M. F. A. Methods for the calculation of titres. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 445—459.
- Dean D. J., Sbelseth M. K., Atanasiu P. The fluorescent antibody test. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 88—95.
- King A. A. Cell culture of rabies virus. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 114—130.
- Laloševi? D., Laloševi? V., Lazarevi?-Ivanc Lj., Kne?evi? I. BHK-21 cell culture rabies vaccine: immunogenicity of a candidate vaccine for humans. In: Towards the Elimination of Rabies in Eurasia. Dev Biol. Basel, Karger, 2008. — 131:421—429.
- Muhammad Zubair Yousaf, Muhammad Qasim, Sadia Zia, Muti ur Rehman Khan, Usman Ali Ashfaq, Sanaullah Khan. Rabies molecular virology, diagnosis, prevention and treatment. — Virology Journal, 2012. — 9:50.
- Reculard P. Cell-culture vaccines for veterinary use. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 314—323.
- Smith J. S., Yager P. A., Baer G. M. A rapid fluorescent focus inhibition test (RFFIT) for determining rabies virus-neutralizing antibody. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 181—192.
- Tordo N. Characteristics and molecular biology of the rabies virus. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 28—51.
- WHO Expert Consultation on Rabies: first report. WHO technical report series 931. Geneva: WHO; 2004 — 121 p.
- World Organisation for Animal Health (OIE) Тerrestrial manual. Rabies; 2013. — 28 p.
Literature
- L'vov D. K., Alimbarova L. M., Al'xovskij S. V. i dr. Medicinskaya Virusologiya / Pod red. L'vova D. K. — M.: MIA, 2008. — 656 c.
- Cucaeva A. A., Mikulinskij Yu. E., Vysekancev I. P. Xolodovoj stress i biologicheskie sistemy / Pod red. Cucaevoj A. A. — K.: Nauk. dumka, 1991. — 176 s.
- Aubert M. F. A. Methods for the calculation of titres. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 445—459.
- Dean D. J., Sbelseth M. K., Atanasiu P. The fluorescent antibody test. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 88—95.
- King A. A. Cell culture of rabies virus. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 114—130.
- Laloševi? D., Laloševi? V., Lazarevi?-Ivanc Lj., Kne?evi? I. BHK-21 cell culture rabies vaccine: immunogenicity of a candidate vaccine for humans. In: Towards the Elimination of Rabies in Eurasia. Dev Biol. Basel, Karger, 2008. — 131:421—429.
- Muhammad Zubair Yousaf, Muhammad Qasim, Sadia Zia, Muti ur Rehman Khan, Usman Ali Ashfaq, Sanaullah Khan. Rabies molecular virology, diagnosis, prevention and treatment. — Virology Journal, 2012. — 9:50.
- Reculard P. Cell-culture vaccines for veterinary use. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 314—323.
- Smith J. S., Yager P. A., Baer G. M. A rapid fluorescent focus inhibition test (RFFIT) for determining rabies virus-neutralizing antibody. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 181—192.
- Tordo N. Characteristics and molecular biology of the rabies virus. In: Laboratory techniques in rabies. Geneva: WHO; 1996. — P. 28—51.
- WHO Expert Consultation on Rabies: first report. WHO technical report series 931. Geneva: WHO; 2004 — 121 p.
- World Organisation for Animal Health (OIE) Тerrestrial manual. Rabies; 2013. — 28 p.